Экспрессия генов. Что такое экспрессия генов?
- Экспрессия генов. Что такое экспрессия генов?
- Регуляция экспрессии генов у прокариот и эукариот. Регуляция экспрессии генов у прокариот
- Репликация ДНК. Процесс репликации ДНК
- Позитивная и негативная Регуляция экспрессии генов. Регуляция экспрессии генов у прокариот
- Оперон
- Регуляция экспрессии генов эукариот на уровне транскрипции. Регуляция экспрессии генов на уровне транскрипции у эукариот
- Общие принципы генетического контроля экспрессии генов
- Экспрессия в медицине. Определение экспрессии
- Метилирование ДНК. Вступление. Открытия лаборатории Белозерского.
Экспрессия генов. Что такое экспрессия генов?
"Начать свое выступление я бы хотел с объяснения некоторых терминов и концепций биологии клетки и клеточной генетики.
В самом центре рисунка 1. находиться элемент, который называется - ядро клетки, и внутри этого клеточного ядра содержится днк - то образование, в котором хранится вся информация о том, что мы есть.
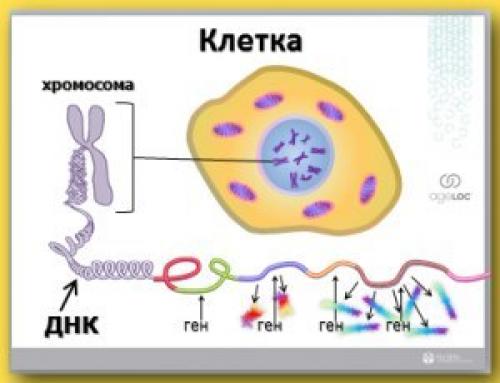 Эти структуры, хранящие информацию о нас, называются хромосомы. И вокруг хромосом, как показано здесь на рисунке, плотно обвиваются наши молекулы днк. И если размотать эту цепочку, то она состоит из отдельных участков - индивидуальных генов. Каждый из этих генов содержит информацию, которая представляет собой отдельный уникальный кирпичик, из которого строится конкретный человек.
Эти структуры, хранящие информацию о нас, называются хромосомы. И вокруг хромосом, как показано здесь на рисунке, плотно обвиваются наши молекулы днк. И если размотать эту цепочку, то она состоит из отдельных участков - индивидуальных генов. Каждый из этих генов содержит информацию, которая представляет собой отдельный уникальный кирпичик, из которого строится конкретный человек.
 Человек, его тело и организм состоят из около 50 триллионов клеток. И каждая клетка полный комплект днк содержит. И в каждом этом комплекте днк находится порядка 20 тысяч генов. Я уже упомянул о том, что каждый ген служит строительным кирпичиком вашего организма. Но возможно сейчас прозвучит термин, с которым вы пока не знакомы, но он очень важен для понимания той науки, которая стоит за созданием аgeLОС.
Человек, его тело и организм состоят из около 50 триллионов клеток. И каждая клетка полный комплект днк содержит. И в каждом этом комплекте днк находится порядка 20 тысяч генов. Я уже упомянул о том, что каждый ген служит строительным кирпичиком вашего организма. Но возможно сейчас прозвучит термин, с которым вы пока не знакомы, но он очень важен для понимания той науки, которая стоит за созданием аgeLОС.
 Что такое экспрессия генов?
Что такое экспрессия генов?
 Каждый из генов нашего организма отвечает за создание определенного продукта, но выработка этого продукта из расчета на один ген - различна. Некоторые из них вырабатывают много того, что должны, другие - недостаточно. И как раз это и определяет степень (концепцию) экспрессии гена. Экспрессия - это скорость (например - поезд - экспресс) в нашем случае - это активность генов. Экспрессия некоторых генов находится на невысоком уровне, а у других - степень экспрессии высокая. Экспрессия генов не измеряется в категориях - хорошая экспрессия или плохая экспрессия, каждый из генов имеет свою собственную экспрессию.
Каждый из генов нашего организма отвечает за создание определенного продукта, но выработка этого продукта из расчета на один ген - различна. Некоторые из них вырабатывают много того, что должны, другие - недостаточно. И как раз это и определяет степень (концепцию) экспрессии гена. Экспрессия - это скорость (например - поезд - экспресс) в нашем случае - это активность генов. Экспрессия некоторых генов находится на невысоком уровне, а у других - степень экспрессии высокая. Экспрессия генов не измеряется в категориях - хорошая экспрессия или плохая экспрессия, каждый из генов имеет свою собственную экспрессию.
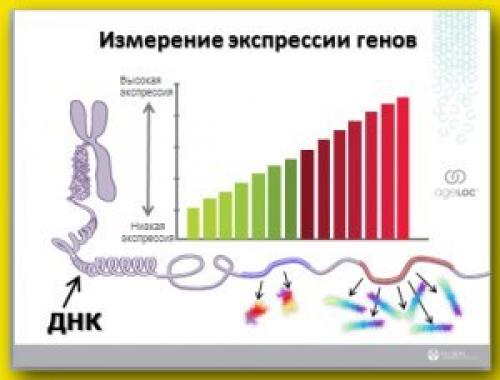
 Генетики - это ученые, которые изучают гены. И их очень интересует возможность измерить величину экспрессии каждого гена. И для этого в научной лаборатории Nuskin совместно с лабораторией Lifegentechnology была разработана специальная цветная шкала, которая позволяет хорошо укладывать в различные цветовые комбинации величину экспрессии генов. Рисунок 2.
Генетики - это ученые, которые изучают гены. И их очень интересует возможность измерить величину экспрессии каждого гена. И для этого в научной лаборатории Nuskin совместно с лабораторией Lifegentechnology была разработана специальная цветная шкала, которая позволяет хорошо укладывать в различные цветовые комбинации величину экспрессии генов. Рисунок 2.
 Лишь в том случае, если экспрессия гена находиться на невысоком уровне, то он окрашивается в один из цветов зеленой части спектра. А если ген имеет высокую экспрессию, то при нанесении на эту цветовую палитру он окрашивается в красной части спектра.
Лишь в том случае, если экспрессия гена находиться на невысоком уровне, то он окрашивается в один из цветов зеленой части спектра. А если ген имеет высокую экспрессию, то при нанесении на эту цветовую палитру он окрашивается в красной части спектра.
 Вы помните, что я упоминал, что в каждой днк находиться около 20 тысяч генов. Это может показаться безумием, но для разработки нашей технологии Ageloc нам приходиться иметь дело со всеми 20 тысячами генов. Не просто каждый ген обладает собственным размером экспрессии, собственной величиной экспрессии, экспрессия наших генов различна в разные периоды жизни человека.
Вы помните, что я упоминал, что в каждой днк находиться около 20 тысяч генов. Это может показаться безумием, но для разработки нашей технологии Ageloc нам приходиться иметь дело со всеми 20 тысячами генов. Не просто каждый ген обладает собственным размером экспрессии, собственной величиной экспрессии, экспрессия наших генов различна в разные периоды жизни человека.
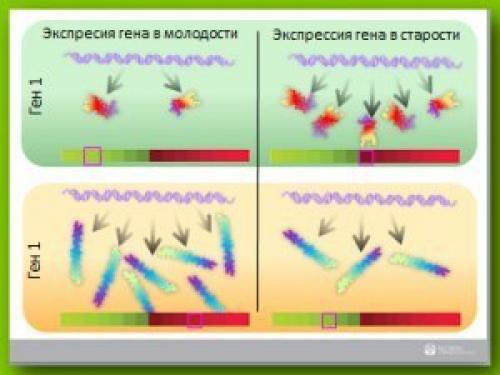
 На слайде (рисунок 3. ) показаны два разных гена, один имеет низкую экспрессию, другой - высокую, и далее мы видим, как сильно меняется величина экспрессии генов, когда возраст человека увеличивается, может получиться прямо противоположная картинка.
На слайде (рисунок 3. ) показаны два разных гена, один имеет низкую экспрессию, другой - высокую, и далее мы видим, как сильно меняется величина экспрессии генов, когда возраст человека увеличивается, может получиться прямо противоположная картинка.
 Ген, который имел высокую экспрессию, когда человек был молод, с течением времени, в результате старения, его экспрессия понижается, а экспрессия гена, который был менее активен в молодости, возрастает.
Ген, который имел высокую экспрессию, когда человек был молод, с течением времени, в результате старения, его экспрессия понижается, а экспрессия гена, который был менее активен в молодости, возрастает.
Таким образом становиться понятно, что важно не просто изучать степени экспрессии генов, велика она или мала, важно обеспечить сохранение того уровня экспрессии генов, который был у человека в молодости. И на этой цветовой палитре очень хорошо видно, что ген, у которого была низкая активность в молодом возрасте, с возрастом повышается, и аналогично, ген, у которого была высокая экспрессия в молодости, с возрастом - снижается, перемещаясь в другую часть спектра.
Это та концепция, которая называется "Концепцией Экспрессии Генов", и она имеет исключительную важность для понимания концепции Ageloc в целом.
А как генетики изучают экспрессию генов и исследуют ее на протяжении нашей жизни?
Для этого был разработан специальный генетический чип (рисунок 4. . и в этом небольшом окошке темного цвета хранится огромное количество информации.
Фактически один - единственный чип способен измерить весь геном человека, все 20 тысяч генов. Каждая из этих точек обозначает один из 20 тысяч генов, которые измеряются данным чипом. И можно использовать такой генетический чип для измерения экспрессии всех генов конкретного человека в разные периоды его жизни, и при этих замерах точки, обозначающие экспрессию отдельных генов, будут в течении жизни изменять свой цвет в зависимости от изменения экспрессии.
Чтобы еще лучше проиллюстрировать вам концепцию экспрессии генов, я хочу рассказать вам совершенно замечательную историю об исследовании экспрессии генов у однояйцевых близнецов.
Эти две близняшки родились в 1908 году (рисунок 5. . этой фотографии почти сто лет, она была сделана, когда девочкам было пять лет. Это однояйцевые близнецы, то есть их молекулы днк идентичны. Они действительно выглядят одинаково? Как вам видится? Согласны, что они абсолютно идентичны, то есть совершенно одинаковы, как внешне, так и внутренне. Теперь давайте посмотрим, как они выглядели в 12 лет. Они по-прежнему выглядят одинаково?
А на фотографии где им 21 год (рисунок 6.? А теперь давайте посмотрим на фотографию, когда им исполнилось по 102 года (рисунок 7. . а сейчас они выглядят одинаково? Их, конечно, можно принять за сестер, они похожи друг на друга, как родственники, но они уже не так похожи, как однояйцевые близнецы.
Как может так получиться, что в старости два организма с идентичными молекулами днк выглядят по-разному? Что у них изменилось? Молекула днк? Нет, днк не меняется никогда. И вот именно здесь вступает в дело замечательная наука, изучающая экспрессию генов.
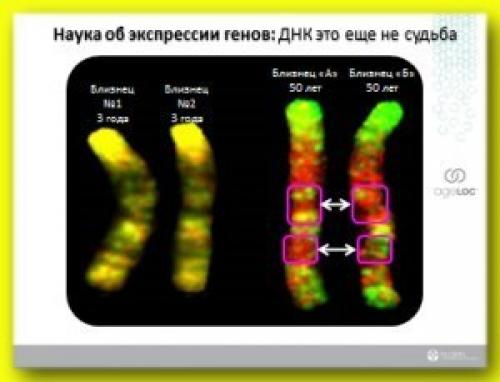
Здесь представлены хромосомы четырех человек (рисунок 8. . правая часть представлена хромосомами близнецов. Желтая часть - это хромосомы близнецов молодого возраста, а красная - это хромосомы близнецов пожилого возраста. Вы помните, что хромосомы состоят из молекул днк и то отличие в цветах, которое мы видим на фотографиях, оно обусловлено именно экспрессией генов. И если посмотреть на окраску хромосом близнецов молодого возраста, маленьких детей, то мы видим, что отличия в цветах практически - нет, потому что отсутствует различие в экспрессии их генов. Но днк близнецов пожилого возраста уже начинают различаться по цвету, потому что в силу разных условий жизни взрослых людей у них по-разному меняется экспрессия генов и идет процесс старения.
Я хотел бы еще раз подчеркнуть, что молекулы днк как молодых близнецов, представленных здесь, так и днк пожилых людей продолжают оставаться абсолютно идентичными друг другу, только экспрессия их генов с возрастом меняется! И если наш возраст влияет на экспрессию генов, то разве неинтересно узнать, какие пищевые компоненты, какие нутриенты могут повлиять положительным образом на то, как будет меняться экспрессия наших генов с возрастом.
Давайте сейчас прервемся в обсуждении и объяснении того, как осуществляется экспрессия наших генов. И позвольте мне представить вам научный совет Pharmanex.
В трех странах мира расположены наши научные лаборатории. Одна из наших лабораторий расположена в Пекине, есть лаборатория в Шанхае, есть лаборатория в штаб-квартире нашей компании, в прово. И не то чтобы у нас было очень много ученых, просто наши ученые имеют очень сильно отличающийся от других багаж знаний, и знания очень разнообразные.
Как же нам помогает такой разнообразный багаж знаний наших ученых?
Ведь у истинных ученых очень часто возникают различные мнения по научным вопросам. Относясь к делу творчески, мы внесли различные научные позиции как элемент научных изысканий, заставляющий подходить к проблеме старения - комплексно.
Мы решили проанализировать идею о том, что все гены нашего организма начинают с возрастом работать по-другому и изучить специфику работы генов в молодом и пожилом возрасте. Но добиться понимания того, как осуществляется экспрессия генов на протяжении всей жизни человека, - это очень сложная задача. И несмотря на то, что у нас более ста ученых, никто из них, взятый отдельно, не в состоянии в полной мере изучить и оценить, как осуществляется экспрессия генов, например, животного, на всем протяжении жизни этого животного.
На самом деле на всей нашей планете есть только одна группа (коллектив, команда) ученых, которая владеет этим знанием.
И вот эта команда ученых из лаборатории "Лайф Ген Технлогия". Я не могу переоценить важность нашего сотрудничества с этой группой ученых. Есть в мире косметические компании, которые стремятся к тому, чтобы положительным образом изменить экспрессию генов, но принципиальное отличие состоит в том, что они стараются воздействовать только на один - два гена из всего генома человека. Но процесс старения - он гораздо более сложный и не укладывается в один или два гена. По мере того, как мы стареем, затрагивается экспрессия тысяч и тысяч генов, и нужно понимать, как можно влиять на этот процесс. И ученые "Лайф Ген Технология", они разработали у себя вот это знание, то как работают все наши гены.
Мы рассказали вам о концепции экспрессии генов и вы знаете, что некоторые гены имеют высокую экспрессию, а некоторые - низкую. И помните цветовую палитру, тот спектр, который я вам показывал? Зеленый означает низкую экспрессию генов, а красный - высокую. Это очень сложно, взять 20 тысяч генов и посмотреть на каждый из них в отдельности.
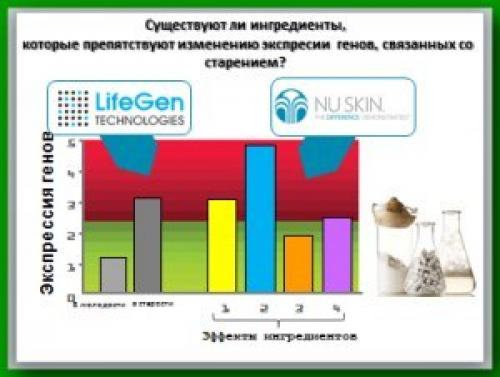
Но нашими учеными разработана методика, которая позволяет упростить задачу, сгруппировав определенное количество генов в некую единую категорию, для которой выводится средняя арифметическая экспрессии. И именно это показано на данном графике (рисунок 9. ), данной диаграмме - это среднее арифметическое экспрессии генов в молодом организме и далее показан среднеарифметический показатель для той же самой группы генов, и видно, насколько сильно экспрессия генов этой группы возросла в пожилом возрасте. И можно наглядно видеть то, как возросла экспрессия этих генов по мере старения организма. И никто в мире не понимает эти процессы лучше, чем ученые из "Лайфгентехнологии".
Но когда речь заходит об ингредиентах и нутриентах, и о том, как они влияют на экспрессию генов, здесь уже приоритет ученым Nuskin. И здесь мы видим, то какое воздействие определенный набор ингредиентов оказывает на эти именно гены.
И мы видим, что ингредиент номер 3 оказался способен уменьшить экспрессию генов данной группы до уровня практически сопоставимым с тем, который естественным образом присутствовал в данном организме в молодом возрасте.
И таким образом мы проанализировали тысячи и тысячи ингредиентов и компонентов, стремясь понять, то, как они воздействуют на экспрессию генов и насколько данный конкретный ингредиент способен сохранить экспрессию данной конкретной группы генов на уровне, присущем молодому организму …" автор: Гленн чейни, отдел исследований и разработок Pharmanex.
Регуляция экспрессии генов у прокариот и эукариот. Регуляция экспрессии генов у прокариот
Переключение генов лучше всего изучено у прокариот (бактерий). Рассмотрим механизмы регуляции активности генов на примере лактозного оперона кишечной палочки (Escherichia coli) – классического объекта генетики микроорганизмов. Единицей регуляции экспрессии генов у прокариот является оперон.
Оперон – это участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т – терминатор. (В состав других оперонов может входить до 10 структурных генов и более.)
Промотор – это регуляторный участок ДНК, который служит для присоединения РНК-полимеразы к молекуле ДНК. В лактозном опероне присоединение РНК-полимеразы происходит с помощью комплекса CAP-цАМФ (CAP – это специфический белок; в свободной форме является неактивным активатором, цАМФ – циклоаденозинмонофосфат – циклическая форма аденозинмонофосфорной кислоты).
Оператор – это регуляторный участок ДНК, который способен присоединять белок-репрессор, который кодируется соответствующим геном lac. Если репрессор присоединен к оператору, то РНК-полимераза не может двигаться вдоль молекулы ДНК и синтезировать мРНК.
Структурные гены кодируют три фермента, необходимые для расщепления лактозы (молочного сахара) на глюкозу и галактозу. Молочный сахар лактоза – менее ценный продукт питания, чем глюкоза, поэтому в присутствии глюкозы сбраживание лактозы является невыгодным для бактерии процессом. Однако при отсутствии глюкозы бактерия вынуждена переходить на питание лактозой, для чего синтезирует соответствующие ферменты Z (β-галактозидазу), Y (галактозидпермеазу), А (тиогалактозидтрансацетилазу).
Терминатор – это регуляторный участок ДНК, который служит для отсоединения РНК-полимеразы после окончания синтеза мРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы.
Для регуляции работы оперона необходим ген cya, кодирующий белок CYA, который катализирует образование цАМФ из АТФ, Если в клетке имеется глюкоза, то белок CYA вступает с ней в реакцию и переходит в неактивную форму. Таким образом, глюкоза блокирует синтез цАМФ и делает невозможным присоединение РНК-полимеразы к промотору. Следовательно, глюкоза является репрессором лактозного оперона.
Если же в клетке имеется лактоза, то она взаимодействует с белком-репрессором и превращает его в неактивную форму. Белок-репрессор, связанный с лактозой, не может присоединиться к оператору и не преграждает путь РНК-полимеразе. Таким образом, лактоза является индуктором лактозного оперона.
Предположим, что первоначально в клетке имеется только глюкоза. Тогда белок-репрессор присоединен к оператору, а РНК-полимераза не может присоединиться к промотору. Оперон не работает, структурные гены выключены.
При появлении в клетке лактозы и при наличии глюкозы белок-репрессор отщепляется от оператора и открывает путь РНК-полимеразе. Однако РНК-полимераза не может присоединиться к промотору, поскольку глюкоза блокирует синтез цАМФ. Оперон по-прежнему не работает, структурные гены выключены.
Если же в клетке имеется только лактоза, то белок-репрессор связывается с лактозой, отщепляется и открывает путь РНК-полимеразе. В отсутствии глюкозы белок CYA катализирует синтез цАМФ, и РНК-полимераза присоединяется к промотору. Структурные гены включаются, РНК-полимераза синтезирует мРНК, с которой транслируются ферменты, обеспечивающие сбраживание лактозы.
Таким образом, лактозный оперон находится под двойным контролем индуктора (лактозы) и репрессора (глюкозы). Общий принцип регуляции активности структурных генов в этих оперонах показан на рис. _____ (см. рисунки в конце лекции).
Общие принципы регуляции активности генов
Кроме лактозного оперона, у кишечной палочки хорошо изучены и другие опероны: триптофановый (trp), гистидиновый (his) и другие. Общий принцип регуляции активности структурных генов в этих оперонах показан на рис. _____ (см. рисунки в конце лекции).
Общие принципы регуляции активности генов в оперонах разработали Франсуа Жакоб и Жак Моно (1961; Нобелевская премия 1965). Согласно концепции Жакоба–Моно, единицей регуляции активности генов у прокариот является оперон. Транскрипция группы структурных генов, регулируется двумя элементами – геном-регулятором и оператором. Оператор часто локализуется между промотором и структурными генами; ген-регулятор может локализоваться рядом с опероном или на некотором расстоянии от него.
Если продуктом гена-регулятора является белок-репрессор, его присоединение к оператору блокирует транскрипцию структурных генов, препятствуя присоединению РНК-полимеразы к специфичному участку – промотору, необходимому для инициации транскрипции. Напротив, если белком-регулятором служит активный апоиндуктор, его присоединение к оператору создает условия для инициации транскрипции. В регуляции работы оперонов участвуют также низкомолекулярные вещества – эффекторы, выступающие как индукторы либо корепрессоры структурных генов, входящих в состав оперонов.
Различают индуцируемые (включаемые) и репрессируемые (выключаемые) опероны в зависимости от типа влияния на их работу молекул-эффекторов.
У индуцируемых оперонов эффектор присоединяется к белку-репрессору и блокирует его связывание с оператором, препятствуя транскрипции структурных генов. Такой тип регуляции работы оперона называют негативным. При негативном контроле эффектор, являющийся корепрессором, присоединяется к неактивному репрессору и активирует его. В результате репрессор приобретает способность присоединяться к оператору и тем самым блокировать транскрипцию оперона. Таким образом, при негативном контроле эффектор связывается с репрессором, что приводит к его инактивации либо активации и соответственно индуцирует либо репрессирует транскрипцию оперона.
Наряду с этим, индуцируемые опероны могут находиться под позитивным контролем регуляции, при котором эффектор связывается с регуляторным белком и активирует его. Активный апоиндуктор присоединяется к оператору, что обеспечивает возможность транскрипции оперона. Оба типа контроля регуляции действуют и в отношении репрессируемых оперонов. При позитивном контроле функционирования репрессируемого оперона корепрессор связывается с активным апоиндуктором. Такой комплекс не может присоединяться к оператору, и структурные гены не транскрибируются. При позитивном контроле эффектор присоединяется не к репрессору, а к апоиндуктору, что разрешает, или, напротив, блокирует транскрипцию в зависимости от того, какую форму (активную или неактивную) приобретает апоиндуктор в результате связывания с эффектором. Поскольку при транскрипции оперона, состоящего из нескольких структурных генов, образуется один общий транскрипт в виде молекулы полицистронной мРНК, все эти гены экспрессируются координировано.
Репликация ДНК. Процесс репликации ДНК
Информация, записанная в ДНК, должна быть не только реализована в процессе развития клеток и организмов, но и в полном объеме передана следующему поколению. С этой целью перед делением клетки в ней осуществляется процесс репликации , т.е. удвоения количества ДНК.
Информация о механизме репликации содержится в самой ДНК: одни гены кодируют ферменты, синтезирующие предшественники ДНК — нуклеотиды, другие — ферменты, обеспечивающие соединение активированных нуклеотидов в единую цепочку. Механизм репликации был впервые постулирован Дж. Уотсоном и Ф. Криком, которые отмечали, что комплементарность цепей ДНК наводит на мысль, что эта молекула может удваивать саму себя. Они предположили, что для удвоения необходим разрыв водородных связей и расхождение цепей, каждая из которых играет роль матрицы при синтезе комплементарной цепи. В результате одного акта удвоения образуются две двунитиевые молекулы ДНК, в каждой из которых имеется одна материнская нить и одна новая (см. рис.).
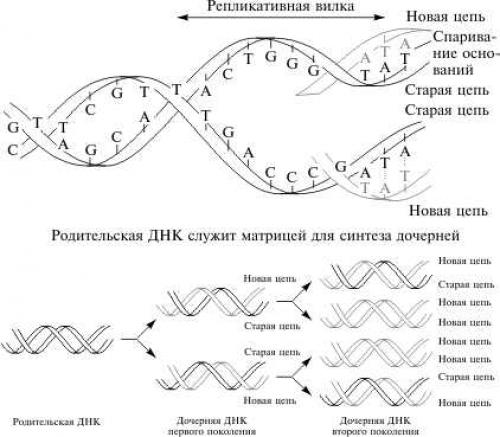
Полуконсервативная репликация ДНК
Механизм получил название полуконсервативной репликации . Позже матричная природа и постулированный принцип репликации ДНК были подтверждены многочисленными экспериментальными данными.
Репликация ДНК начинается в специфических точках хромосомы — сайтах инициации репликации (origin). Процесс репликации обслуживается большим количеством ферментов. Наиболее полно изучен аппарат репликации бактериальной ДНК, особенно E. coli. Функцию расплетания молекулы ДНК у прокариот выполняют специфические ферменты геликазы , которые используют для работы энергию гидролиза АТФ до АДФ. Они часто функционируют в составе белкового комплекса, осуществляющего перемещение вилки и репликацию расплетенных нитей. Удерживают нити ДНК от воссоединения другие специфические белки, связывающиеся с одноцепочечными участками. Эти участки, разошедшиеся в разные стороны, образуют характерную структуру — репликативную вилку (вилку Кернса). Это — та часть молекулы ДНК, в которой в данный момент осуществляется синтез новой цепи. В продвижении вилки большую роль играет белок гираза , относящийся к разряду топологических изомераз. Он обнаружен только у бактерий. Гираза — это релаксирующий фермент, который, производя двунитиевые разрывы, снимает положительные (перед вилкой) и способствует образованию отрицательных (сзади вилки) супервитков в релаксированной ДНК.
Позитивная и негативная Регуляция экспрессии генов. Регуляция экспрессии генов у прокариот
У прокариот пока молекула РНК синтезируется на участке ДНК, она тут же может транслироваться (начиная с уже синтезированного конца). Поэтому у них регуляция экспрессии (активности) генов осуществляется почти исключительно на уровне ДНК, так как в РНК часто невозможно внести какие-нибудь изменения до ее трансляции.
В 1961 г. Жакобом и Моно была предложена модель оперона как системы регуляции генов у бактерий. Оперон состоит из промотора, оператора, структурных генов оперона (их может быть разное количество) и терминатора. В области промотора прикрепляется фермент РНК-полимераза. В области оператора присоединяется белок-репрессор, который кодируется отдельно отстоящим от оперона геном-регулятором (может быть сцеплен со своим опероном, а может находиться на расстоянии).
Если белок-репрессор соединяется с оператором, то транскрипция всех структурных генов оперона становится невозможной, так как РНК-полимераза не может перемещаться по цепи ДНК.
В свою очередь активность белка-репрессора может блокироваться определенным для него низкомолекулярным соединением — индуктором (тем или иным питательным веществом бактерий). В результате взаимодействия с индуктором белок-репрессор видоизменяется и уже не может присоединиться к оператору своего оперона. В этом случае гены оперона экспрессируются (т. е. на них идет синтез).
Бывает обратная ситуация, когда индуктор активирует белок-репрессор.
Таким образом, в зависимости от того, какие индукторы находятся в цитоплазме, у прокариот экспрессируются те или иные генные группы.
Вышеописанный механизм экспрессии генов относится к негативной регуляции, так как гены транскрибируются, если они не выключены репрессором. И наоборот: не транскрибируются, если выключены.
Кроме негативной регуляции у бактерий существует также позитивная. В этом случае вместо белка-репрессора действие оказывает белок-активатор. На эти белки также действуют индукторы, активируя или инактивируя их.
Также у прокариот были выявлены опероны, которые актируются двумя регуляторными белками, соединенными друг с другом.
Оперон
Оперон — функциональная единица генома у прокариот , в состав которой входят цистроны ( гены , единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами . Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов).
Характерным примером оперонной организации генома прокариот является лактозный оперон , триптофановый , пиримидиновый и bgl опероны у Escherichia coli
Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор.
Регуляция экспрессии генов эукариот на уровне транскрипции. Регуляция экспрессии генов на уровне транскрипции у эукариот
Несмотря на то, что основные принципы регуляции транскрипции генов у прокариотических и эукариотических организмов остаются неизменными - через специфические взаимодействия белков и нуклеиновых кислот друг с другом, а также между собой, данный процесс у эукариот характеризуется рядом существенных особенностей. Это связано, прежде всего, с необходимостью поддержания координированной экспрессии эукариотических генов в более сложноорганизованной генетической системе. Достаточно вспомнить, что в организме человека гистологически различают, по крайней мере, 100 типов клеток, формирующих его органы и ткани. Для любого типа клеток характерен свой уникальный набор экспрессирующихся генов, которые начинают функционировать во время дифференцировки клеток-предшественников. Кроме того, сам процесс формирования органов и тканей сопровождается пролиферацией строго определенных групп клеток, а также упорядоченным во времени и пространстве перемещением клеток. Все эти особенности жизнедеятельности клеток высших организмов обеспечиваются функционированием их генов.
Общие принципы генетического контроля экспрессии генов
Важнейшим фактором регуляции генной активности являются элементы генома, отвечающие за синтез регуляторных
Рис. 3.84. Схема конститутивных и регулируемых генов.
А — конститутивный ген; Б, В — регулируемые гены; Б,, Б2— связывание РНК-полимеразы с промотором возможно лишь в отсутствие белка-репрессора, который специфически соединяется с оператором, частично или полностью перекрывающим промоторную последовательность; В — связывание РНК-полимеразы с промотором облегчается белком-активатором (апоиндуктором), который специфически узнает область оператора, расположенную перед промотором; стрелкой обозначено направление транскрипции
белков, — гены-регуляторы. Соединяясь с определенными нуклеотидными последовательностями ДНК, предшествующими структурной части регулируемого гена — операторами, белки-регуляторы способствуют или препятствуют соединению РНК-полимеразы с промотором. Если белок-регулятор взаимодействует с оператором, занимающим часть промотора или расположенным между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соединиться с промоторной последовательностью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществляется негативный контроль экспрессии гена со стороны гена-регулятора (рис. 3.85). Если промотор обладает слабой способностью соединяться с РНК-полимеразой, а ему предшествует область, узнаваемая белком-регулятором, присоединение последнего непосредственно перед промотором к молекуле ДНК облегчает связывание РНК-полимеразы с промотором, вслед за чем следует транскрипция. Такие белки называют активаторами (или апоиндукторами) , а контроль экспрессии гена со стороны гена-регулятора — позитивным (см. рис. 3.85).
Экспрессия в медицине. Определение экспрессии
Что такое экспрессия? В медицине это генетический процесс, посредством которого содержащаяся в генах информация преобразуется в определенные клеточные структуры. Экспрессивность в биологии – это неразрывно связанное с жизнедеятельностью и здоровьем человека определение, которое получает дальнейшее развитие в психологии и психотерапии. К этой же медицинско-биологической сфере относится понятие «экспрессировать» – это означает транскрибировать и транслировать ген.
Экспрессивность – это степень выраженности определенного состояния, чувства, эмоции, отношения. Понятия «экспрессивность» и «экспрессия» применяются не только в психологии, но и в театроведении, искусствоведении, если требуется подчеркивать уровень выраженности духовности. А также показать средства выражения, к примеру, живопись, музыка, архитектурные элементы. Получается, что в имеющихся определениях данных терминов есть указания на связь этого явления с душевным и духовным миром людей.
Экспрессия – это выразительность чувств и эмоций, показ внутреннего состояния путем проявления его во внешности.
Противоположным по смыслу значением является импрессивность – склонность к внутренним, а не внешним переживаниям, накоплению преимущественно отрицательных чувств. О таких людях говорят, что они держат переживания в себе, не склонны делиться эмоциями и проблемами, обиды и расстройства к ним приходят не сразу, не моментально.
Мнения о связи внутреннего мира людей и экспрессии, которые сформировались по большей части в искусствоведении и философии, дополняются психологией. Сущность этой связи в психологическом выражении видна в том, что экспрессия получает место не только сопровождения действий внешне. Она выступает частью данных явлений, формой их проявления.

Экспрессивная девушка рассказывает что-то своей подруге
По этой причине стоит говорить об экспрессии в виде личностного образования , способа познания духовного мира личности, а также в качестве внешнего «Я».
Метилирование ДНК. Вступление. Открытия лаборатории Белозерского.
В конце 1960-х годов в МГУ, на одном из семинаров межфакультетской лаборатории биоорганической химии, сотрудник А.Н. Белозерского , Борис Ванюшин, занимавшийся изучением необычных нуклеотидов в составе ДНК, высказал идею о том, что некоторые из этих необычных нуклеотидов, а именно 5-метилцитозин, играют важную роль в механизмах регуляции экспрессии генов, клеточной дифференцировки и процессов старения организма.
Образование этого 5-метилцитозина в ДНК было обусловлено ферментативным метилированием обычного цитозина, то есть посадкой на него метильных групп. ДНК в месте посадки на её цитозин метильных групп несколько изменяла свои свойства.
Количество 5-метилцитозина в ДНК менялось в разные периоды жизни организма от максимального, при рождении, до минимального, у дряхлого от старости. Так впервые в СССР была сформирована идея о том, что старение на уровне генома регулируется эпигенетически ( ) – то есть путём механизма вызывающего длительное и стабильное изменение экспрессии генов, которое не связано с изменениями в последовательности ДНК. В последующем Ванюшин опубликовал цикл работ под названием «Метилирование ДНК – эпигенетический контроль за генетическими функциями организма» и в 1977 году вместе с Г. Бердышевым книгу “Молекулярно-генетические механизмы старения”.
К сожалению, из-за сложности и затратности, используемой в те годы технологии изучения процессов метилирования ДНК, дальнейшие исследования в этом направлении были резко сужены.
Сегодня имеется множество разных методов определения метилирования ДНК, некоторые из которых (прохождение цепи ДНК через микропору генноинженерного белка порина А) позволяют читать результаты, не разрушая при этом ДНК, что намного упростило и удешевило подобные исследования ()
Ванюшин был поражен ярко выраженным глобальным уменьшением степени метилирования ДНК во всех органах нерестящейся горбуши, которая после нереста очень быстро погибает от внезапно наступившей старости. Аналогичное понижение уровня метилирования ДНК и запрограммированная гибель наблюдалась и у таких растений как бамбук и агава – процессы, которые запускались только после цветения этих растений. Срубив цветоносный побег можно надолго остановить этот процесс старения и предотвратить резкое понижение уровня метилирования ДНК (). Быстрое снижение общего уровня метилирования ДНК наблюдалось при старении однолетнего растения Arabidopsis thaliana (), но не в случае, когда оно из-за засоленной почвы превращалось в многолетник (). Более того была найдена мутация drd1-6, которая воздействуя на механизм регулирующий старение листьев с помощью RdDM (RNA-directed DNA methylation) (), т.е. через микроРНК воздействующую на метилирование ДНК, вызывала резкое замедление старения листьев Arabidopsis ().